广义忍冬科Caprifoliaceaes s.l.隶属桔梗类川续断目,包含锦带花亚科Diervilloideae、忍冬亚科Caprifolioidea、川续断亚科Dipsacoideae、缬草亚科Valerianoideae、六道木属Zabelia (Rehder) Makino、刺参亚科Morinoideae和北极花亚科Linnaeoideae七大支。全世界约41属,960种,主要分布于温带和寒带地区,部分忍冬科植物具有重要的观赏和药用价值。然而,以往的广义忍冬科系统学研究多基于少数样本的若干叶绿体或核基因片段,亚科之间的系统发育关系存在争议,部分类群如六道木属Zabelia (Rehder) Makino和七子花属Heptacodium Rehd.的系统学位置仍不明晰。

图1. 封面展示广义忍冬科物种花的多样性
按从上到下、由左至右的顺序,封面图片示以下植物:细瘦糯米条Abelia forrestii (Linnaeoideae) (Photograph by S. Landrein ), 猬实Kolkwitzia amabilis (Linnaeoideae) (Photograph by G.M. Li, ), 菰腺忍冬Lonicera hypoglauca (Caprifolioideae) (Photograph by H.X. Wang ), 醉鱼草状六道木Zabelia triflora (Photograph by S. Landrein), Vesalea floribunda (Linnaeoideae) (Photograph by S. Landrein) , 全叶六道木Zabelia integrifolia (Photograph by S. Landrein), Scabiosa comosa (Dipsacoideae) (Photograph by J.L. Zhang ), 柔垂缬草Valeriana flaccidissima (Valerianoideae) (Photograph by S.H. Xia) , 匙叶翼首花 Pterocephalus hookeri (Dipsacoideae) (Photograph by X.X. Zhu ), 大花刺参Acanthocalyx nepalensis subsp. Delavayi (Morinoideae) (Photograph by H.X. Wang ), 海仙花Weigela coraeensis (Diervilloideae) (Photograph by Y.C. Xu ).
为了阐明广义忍冬科亚科之间的系统关系及澄清部分类群的系统学位置,海南大学热带作物学院的王华锋课题组联合美国史密森研究院自然历史博物馆Jun Wen研究员、美国欧柏林大学生物学系Michael J. Moore教授和美国明尼苏达大学Diego F. Morales-Briones博士对广义忍冬科的67个代表性样品进行了杂交富集测序(Hybridization target enrichment sequencing, Hyb-seq),获得来自367个不同基因的707个外显子,96,479个简约信息位点,比对长度为343,609 bp,最短基因座长度为150 bp,最大长度为3,503 bp,平均长度为486 bp。
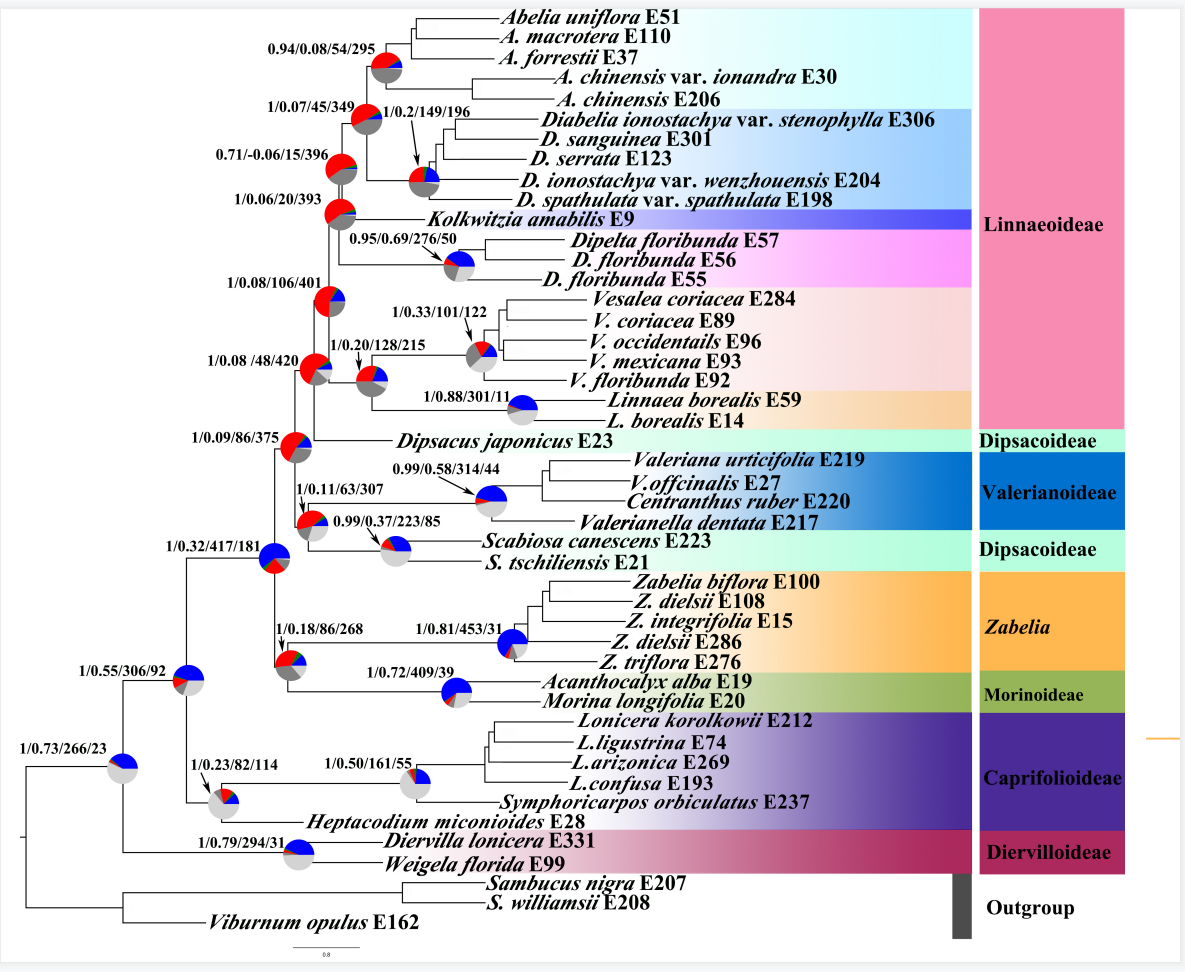
图2 基于核基因数据集,利用ASTRAL-Ⅲ构建的物种树
节点旁边的饼图蓝色部分显示了支持该分支的一致树的比例,绿色部分表示该分支的主要可选拓扑的不一致基因树的比例(绿色),红色部分表示不一致树的比例,深灰色代表无信息基因树的比例(bootstrap支持度<50%),浅灰色代表缺失数据的比例。分支上的数字表示(LPP)/ICA得分/一致基因树数量/所有不一致基因树数量。广义忍冬科主要分支用不同颜色表示,以示区分。
系统发育分析表明广义忍冬科进一步支持可分为7大支:锦带花亚科、忍冬亚科、 北极花亚科、六道木属、刺参亚科、缬草亚科、川续断亚科。在核基因物种树中,六道木属与川续断亚科形成了姊妹关系(PP=1),它们两支共同与除忍冬亚科和锦带花亚科之外的其他所有广义忍冬科的亚科类群构成了姊妹关系,并获得强烈支持(PP=1)。核基因联合系统发育树恢复了川续断亚科的单系,并与缬草亚科构成姊妹关系,得到强烈支持(BS=100),其他分支拓扑结构与物种系统发育树结构类似。在基于叶绿体基因联合构建的系统发育树中,六道木属与刺参亚科共同形成的分支为北极花亚科的姊妹类群(BS=100),双六道木属(Diabelia Landrein)与双盾木属(Dipelta Maxim.)共同形成的分支与猬实属(Kolkwitzia Graebn.)构成姊妹关系(BS=100),这与核基因系统发育树拓扑结构不一致。
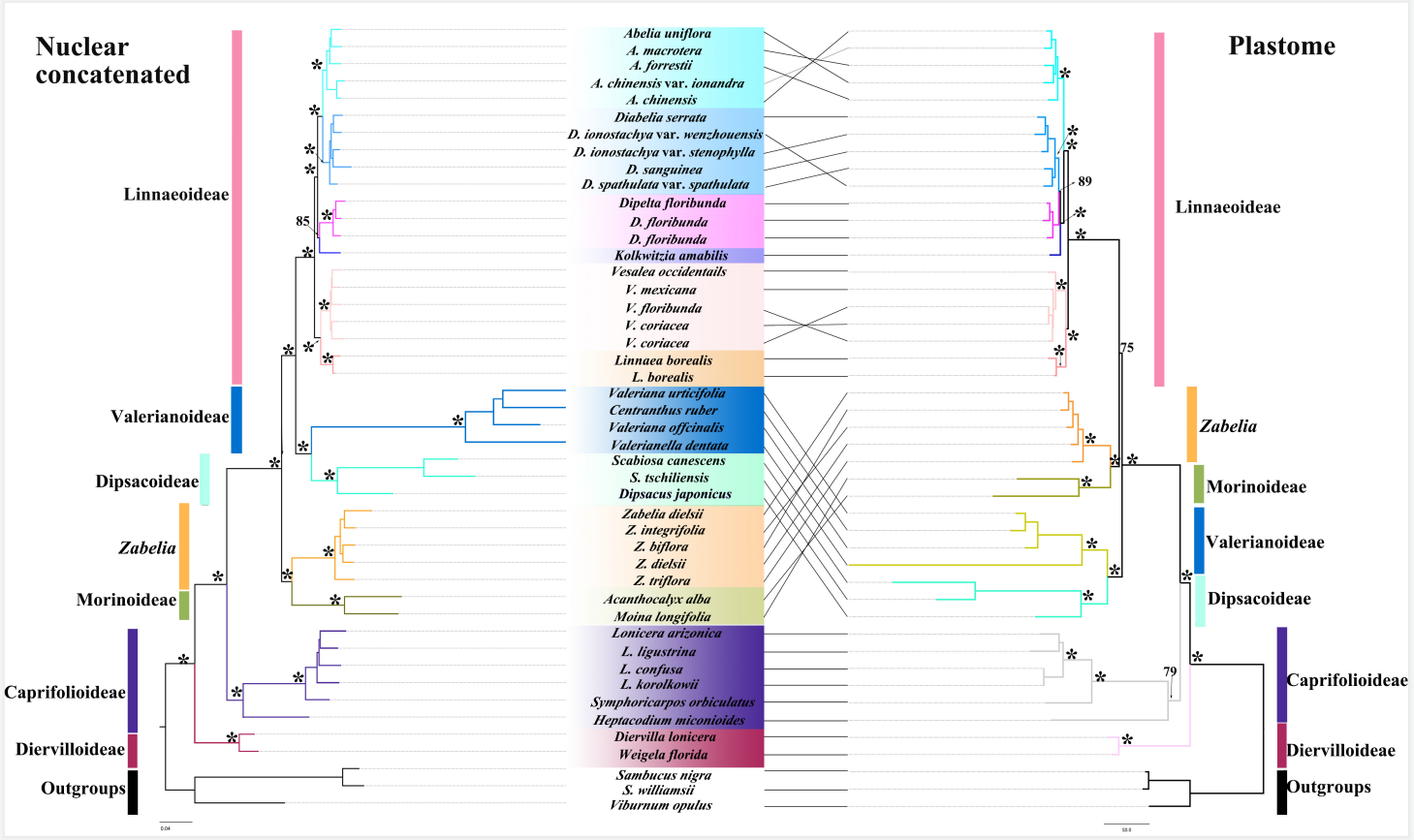
图3 核基因串联树(左)与叶绿体基因串联树(右)的对照图
虚线连接的是两个拓扑结构中相同的物种。在系统发育树分支上的是最大似然引导支持值。星号表示最大的似然引导支持值。广义忍冬科的主要分支用不同颜色表示,以示区分。
为了解析这种不一致,我们对有冲突的物种或支系进行了杂交网状进化分析。结果表明,存在系统发育关系冲突的物种大多和其相邻的支系具有一定的基因流。因此,推测远古杂交可能是造成不同数据集或分析方法所导致的系统发育关系存在冲突的主要原因。
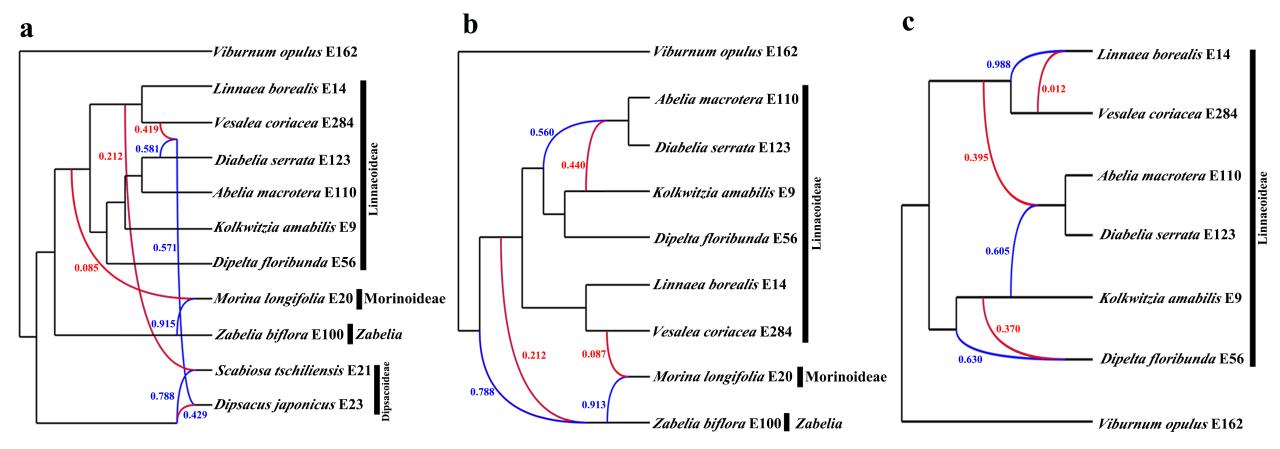
图4 基于不同物种数量的最佳物种网状进化分析
a,11个物种;b,9个物种;c,7个物种。蓝色和红色线旁的数字为遗传概率。红色线表示较小的遗传贡献一方(遗传贡献<0.50)。
此外,基于核基因数据估算广义忍冬科共祖时间大约在白垩纪晚期,约为66.65 Ma (95% HPD = 56.31–69.44 Ma) ,忍冬亚科物种分化出来的时间约为56.35 Ma (95% HPD = 47.76–61.45 Ma),缬草亚科与川续断亚科的分化时间以及北极花亚科物种的共祖时间分别为 41.84 Ma (95% HPD = 34.44–49.35 Ma)和38.32 Ma (95% HPD = 32.66–57.81 Ma)。而刺参亚科与六道木属的分化时间约为33.63 Ma (95% HPD = 32.10–39.89 Ma),这一时期与青藏高原快速隆起时间相重叠,高海拔的地形起到隔离分化作用,同时异质化的生态位也可能促进了分布于该区域广义忍冬科内物种的辐射分化。另外,根据分歧时间估算分析,结合古气候学证据,我们推测古新世晚期和始新世早期地球温度的升高可能迫使广义忍冬科物种向更高的海拔或纬度移动,它们的分布地可能变得破碎,种群之间随之产生隔离分化。基于全叶绿体基因组数据估算的广义忍冬科共祖时间白垩纪晚期到始新世,约为76.43 Ma (95% HPD = 64.81–82.10 Ma),属间的多样性分化可以追溯到渐新世及以后。同时我们发现基于全叶绿体数据比基于核基因数据估算的分化时间更加古老。
最后,通过对广义忍冬科物种的雄蕊数量、花柱是否超过花冠、果实类型、种子和心皮的数量以及是否有副萼6种关键性状进行祖先重建分析。结果表明,其祖先物种的果实类型、花柱是否超过花冠、种子数量和是否有副萼的形态特征是不明确的,而一枚种子和三个心皮可能是该科的祖先物种特征,我们推断川续断属、刺参属和川续断属物种的副萼可能为独立起源。
综上所述,基于叶绿体和核基因组数据的系统发育关系、物种分化时间、形态学和地理分布等综合证据,结合青藏高原隆起造成的隔离造成异质化的生态位可能促进了分布于该区域广义忍冬科内物种的辐射分化,六道木属与近缘类群的系统发育关系、形态特征(例如,六道木属的花期4–6月,花冠成管状,雄蕊不伸出花冠等独特的形态学特征)以及地理分布等与广义忍冬科其他类群都具有显著的不同。因此,我们建议将六道木属提升为广义忍冬科六道木亚科(Zabelioideae)。
本研究成果以A phylogenomic perspective on gene tree conflict and character evolution in Caprifoliaceae using target enrichment data, with Zabelioideae recognized as a new subfamily 为题作为封面研究论文发表于Journal of Systematics and Evolution 2021年第5期。海南大学博士生王宏信和美国明尼苏达大学Diego F. Morales-Briones博士为本文的共同第一作者,王华锋为通讯作者,本研究得到了国家自然科学基金(31660055)的资助,感谢史密森学会国家自然历史博物馆分析生物学实验室的工作人员的支持和帮助。
原文链接https://www.jse.ac.cn/EN/10.1111/jse.1274
 公安部备案号:46010802000190 Copyright © 2024 hainan university
公安部备案号:46010802000190 Copyright © 2024 hainan university





